Cephalotus follicularis
(d'après le fascicule de Jean-Daniel Degreef édité par Dionée)

Remerciements
Publication et origine du nom
Aire géographique
Taxinomie et origine de l'espèce
Evolution des ascidies
Biotope
Description générale
Micro-Anatomie
Physiologie
Culture
Bibliographie
Postscriptum sur l'évolution des ascidies
Je tiens à remercier pour leur aide: Mme KEVERS et M. GASPAR de CEDEVIT qui m'ont fourni de nombreuses
plantules ;
Daniel MORENO de l'Association DIONEE qui m'a apporté une inflorescence;
le Pr. Pierre JOLIVET qui m'a fourni des échantillons de Caltha dionaeifolia
et de la documentation; Holger HENNERN qui m'a envoyé des monstruosités d'un type rarissime; Yves-André UTZ et Isamu KUSAKABE qui m'ont permis de reproduire ici leurs
remarquables dias; M. Marcel LECOUFLE Pierre SIBILLE Peter CATRYSSE qui m'ont fourni de la documentation; les bibliothécaires du Jardin Botanique National de Belgique et de l'Institut de Botanique de l'Université de Liège.
;
Daniel MORENO de l'Association DIONEE qui m'a apporté une inflorescence;
le Pr. Pierre JOLIVET qui m'a fourni des échantillons de Caltha dionaeifolia
et de la documentation; Holger HENNERN qui m'a envoyé des monstruosités d'un type rarissime; Yves-André UTZ et Isamu KUSAKABE qui m'ont permis de reproduire ici leurs
remarquables dias; M. Marcel LECOUFLE Pierre SIBILLE Peter CATRYSSE qui m'ont fourni de la documentation; les bibliothécaires du Jardin Botanique National de Belgique et de l'Institut de Botanique de l'Université de Liège.
|
|
Avril 1990
|
PUBLICATION ET ORIGINE DU NOM
|
Bien que Cephalotus follicularis ait été découvert dès 1791 par le botaniste Archibald MENZIES (oui celui de Drosera et Utricularia menziesii! ) il ne fut publié qu'en 1806 par Jacques Julien Houton de (1755-1834).
On sait que les botanistes attachent une grande importance à l'anatomie des fleurs. C'est ce qu'ils regardent en premier chez une plante. La raison en est qu'au cours de l'évolution la structure
florale varie beaucoup moins rapidement que celle des parties végétatives.
On peut donc grâce aux fleurs espérer saisir des rapports de parenté entre des groupes de végétaux qui n'ont plus grand chose en commun du point de vue feuilles. Il ne faut dès lors pas s'étonner que LABILLARDIERE ait donné à l'
espèce
qui nous occupe un nom faisant allusion à un obscur détail de la structure florale malgré le caractère
frappant des pièges!
Ce détail qui avait frappé LABILLARDIERE est le curieux élargissement du tissu rattachant les
anthères
des étamines entre elles et au filet (le connectif). Les
étamines
possèdent donc une espèce
de petite "tête" de là le nom "Cephalotus" forme latinisée du Grec képhalôtos doté de tête(s). Le nom spécifique follicularis est une allusion aux follicules (petits sacs) c'est à dire
les ascidies qui font tout l'intérêt de l'
espèce
.
| Cephalotus follicularis est sans doute la plante la plus extraordinaire de la flore pourtant peu banale du Sud-Ouest de l'Australie. Il est confiné dans une petite zone de 400 sur (maximum) 80 km.
Celle-ci s'étend d'un petit site près de Busselton au moins jusqu'à la Pallinup River.
Les meilleures localités se trouvent entre Albany et la Frankland River mais celles près d'Albany étaient déjà fort menacées en 1961.
L'espèce
est rare à plus de 16 km de la côte. |
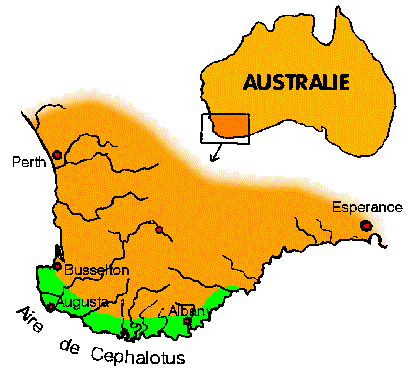 |
Or LABILLARDIERE a réalisé ses collectes de plantes à 200 km à l'Est de cette zone à Esperance Bay. On a donc pensé que l'aire géographique de Cephalotus s'était rétrécie depuis lors.
Une étude récente (CARR & CARR 1976) a montré qu'il n'en était rien.
LABILLARDIERE était le botaniste de la deuxième expédition française en Australie celle conduite par Bruny d'ENTRECASTEAUX en 1791-94. La première expédition avait été commandée par LAPEROUSE (1785-88) la
troisième (1800-04) par Nicolas BAUDIN.
Cette dernière fut marquée par de grosses dissensions politiques liées à la révolution française. A la mort de Louis XVI les botanistes se firent débarquer à Java. Puis le capitaine BAUDIN mourut et le navire fut capturé
par les Anglais avec ses collections végétales. Assez curieusement (vu les circonstances) celles-ci furent rendues à la France et c'est à partir de feuilles d'herbier constituées par le
botaniste LESCHENAULT que LABILLARDIERE réalisa sa publication de
Cephalotus sans jamais avoir été dans les régions où croît cette espèce.
Une feuille d'herbier conservée à Genève porte l'inscription:
| "de Leschenault de la Tour Expédition du Capitaine Baudin ordonnée en l'an 8 en Nouvelle Hollande" |
et de la main de LABILLARDIERE:
|
"Cephalotus follicularis port du roi Georges côte Sud Ouest Nlle holl. 1803".
|
TAXINOMIE ET ORIGINE DE L'ESPECE
|
Le Cephalotus est une plante bizarre qu'il est difficile de rattacher à une
famille végétale
moderne. Les idées concernant sa position taxinomique sont résumées dans les tableaux ci-joints (pour les spécialistes et pour les courageux).
Le gigantesque Ordre Rosales des auteurs anciens a maintenant acquis le statut d'un super-ordre composé d'une série d'ordres parmi lesquels se trouve celui des Saxifragales.
C'est dans ce dernier qu'on place Cephalotus. Comme il n'est pas
possible d'introduire cette
espèce dans une
famille
connue elle en forme une a elle toute seule: les Céphalotacées
Classe
|
Ordres
|
Familles
|
Genres
|
| Choripetalae |
Violales |
Dioncophyllaceae |
Triphyophyllum |
| Saxifragales |
Byblidaceae |
Byblis |
| Cephalotaceae |
Cephalotus |
| Nepenthales |
Nepenthaceae |
Nepenthes |
| Droseraceae |
Aldrovanda |
| Dionaea |
| Drosophyllum |
| Drosera |
| Sarraceniales |
Sarraceniaceae |
Darlingtonia |
| Heliamphora |
| Sarracenia |
|
d'après "Letts Guide to Carnivorous Plants
of the World" de Gordon Cheers
|
L'étude de la répartition des familles à
l'intérieur des Saxifragales et des ordres voisins montre que les ancêtres des Céphalotacées devaient être originaires d'Afrique.
Ils ont du émigrer vers l'Inde et l'Australie avant la formation de l'Océan Indien c'est à dire au Crétacé moyen (vers 100-90 millions d'années avant le présent.
On pense souvent que les urnes de Cephalotus dérivent de feuilles semblables à celles que produit la plante en automne. L'étude micro-anatomique de ces dernières démontre qu'il n'en est rien. Le
pétiole et le
limbe
possèdent des faisceaux homologues de ceux qui dans l'ascidie se rendent vers l'
opercule.
Les feuilles d'hiver ne sont donc pas des feuilles "normales" mais des
ascidies
ratées n'ayant pu se développer suite à des conditions défavorables (développement anormal tératologique). Des feuilles
tératologique plus développées ne sont d'ailleurs pas rares dans
l'espèce qui nous occupe.
On y voit la résurgence de caractères archaïques qui aident grandement à reconstituer
l'évolution du piège.
Celui-ci semble dériver de feuilles peltées divisées
en au moins 4 folioles et quelque peu analogues à celles du marronnier. L'une d'elles produisit cavité de l'urne les trois autres le couvercle.
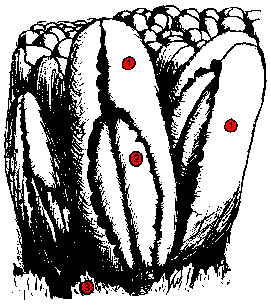 Caltha dionaeifolia
Caltha dionaeifolia
Disposition naturelle des feuilles. Les faces exposées au vent sont dépourvues de stomates (ces derniers sont concentrés sur les faces abritées).
1. surface externe (abaxiale)
2. lobes basaux
3. stipules

Feuille étalée montrant les deux lobes internes qui sont des appendices des deux segments du
limbe repliés vers le haut.
|
La présence de poils sur de nombreuses parties (même souterraines !) de Cephalotus et la structure assez xérophile de son bois sont assez étonnantes.
L'espèce
croit en effet dans des sites qui ne se dessèchent guère. On peut y voir une ancienne adaptation à des marais côtiers battus par le vent et par la pluie qui avec les
inondations périodiques lessivent les feuilles leur enlevant leurs minéraux.
Les poils s'opposent jusqu'à un certain point à ce phénomène. Cette caractéristique et la forme fréquemment aplatie des structures tératologiques rapproche les
ascidies
de Cephalotus des feuilles dites diplophylles décrites par TROLL (1932b). Je ne pense pas que
les urnes de Cephalotus dérivent de telles feuilles mais plutôt de structures
peltées et divisées.
Ce que les feuilles diplophylles apportent c'est de montrer comment ces structures (possédant des
pétioles
analogues à ceux des feuilles peltées
) peuvent se replier au lieu d'être largement déployées comme
des feuilles de marronnier. Ceci pourrait protéger contre le lessivage. Le plus beau cas de diplophyllie est celui de Caltha dionaeifolia. Ceux qui ont assisté à l'A.G. de DIONEE d'Octobre 1989 ont pu observer des feuilles de cette
espèce
apportées par le Pr. P. JOLIVET. Le
genre
Caltha fait partie de la famille
des Renonculacées.
Il y a aussi des feuilles diplophylles dans le genre Alchemilla.
Or ces deux classes de végétaux sont taxinomiquement assez proches des Céphalotacées comme on s'en rendra compte en réexaminant le tableau taxinomique. |
Il y a donc un faisceau de présomptions qui nous mène vers le stade de départ suivant: une feuille
peltée
divisée en quatre folioles les trois qui forment le futur couvercle étant appliquées contre la quatrième (qui se creusera pour donner l'urne). Ceci n'est pas encore un piège à insectes évidemment.
Pourquoi Cephalotus a-t-il évolué vers
la carnivorie et non des espèces comme
Caltha Dionaeifolia et Alchemilla diplophylla qui
ont une structure de base similaire et vivent sur des terrains stériles entourées de plantes
carnivores?
|
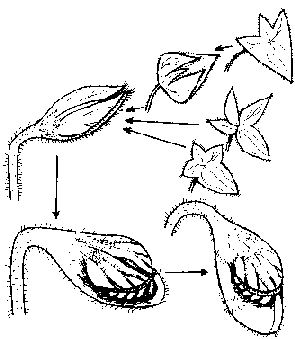
Hypothèse évolutive des Céphalotacées
|
La raison en est peut-être la facilité avec laquelle les végétaux de l'Ordre des Saxifragales sont capables de développer des modifications du bord du
limbe
foliaire. Celui-ci peut devenir pileux garni de dents épaissi en bourrelet et même pourvu de rangées de glandes. Le développement d'une bande de poils dirigés vers la
cavité de la feuille primitive et garnissant les bords des folioles aurait été
un pas important dans l'évolution vers la carnivorie. Un tel dispositif aurait permis l'entrée mais non la sortie des insectes.
De plus il y a des raisons de penser que cette lignée possédait un mécanisme primitif de capture même avant d'avoir développé des pièges à proprement parler.
On est frappé en effet par le fait que toutes les parties de la plante sont couvertes de
poils parmi lesquels se trouvent de petites glandes à nectar. Pourquoi un tel mélange de structures répulsives (les poils) et attractives (les glandes) ? Ces dernières ne
devaient-elles pas encourager les insectes à s'enfoncer dans la toison qui couvrait
la plante quitte à s'y emmêler et à y périr ? Une partie de leurs produits de putréfaction aurait pu être résorbée par les glandes ou même par les poils. HAMILTON (p.39) a décrit la propriété qu'ont certains
trichomes
du rebord du couvercle de capter les colorants. Il en est de même des longs poils situés ailleurs.
En tout cas il est facile d'expliquer les stades ultérieurs de l'évolution d'un piège tel que décrit ici (une feuille repliée aux bords défendus par une bande de poils).
Cette dernière s'est transformée en épiderme à écailles (qui sont des
trichomes très courts! ).
C'est la zone glissante de la collerette du couvercle et de l'entonnoir de
l'ascidie.
Les glandes de Cephalotus dérivent clairement de stomates. Certaines d'entre elles situées à l'intérieur de l'urne ressemblent encore très fort à des stomates.
Les variantes les plus simples des glandes à nectar ont deux cellules superficielles délimitant parfois ce qui pourrait être un vestige de pore. Je me suis aperçu par ailleurs que du bleu de méthylène concentré déposé sur le
limbe
des feuilles lancéolées pénètre occasionnellement dans
les chambres aériennes sous-stomatales! De là il diffuse dans les parois cellulaires de l'épiderme (et sans doute du parenchyme). Cela prouve que la cuticule tapissant les espaces aériens
associés aux stomates est imparfaite. Dans les feuilles primitives
elle a pu permettre l'absorption ou la sortie de substances ce qui a pu aboutir à une spécialisation glandulaire. Ajoutons que les stomates excréteurs d'eau (hydathodes) ne sont pas rares chez les Saxifragacées - autre
famille
de l'ordre Saxifragales - (Saxifraga Zahlbrucknera Chrysosplenium Tellima Mitella Heuchera.
 |
Les aréoles transparentes de l'opercule
avaient sans doute beaucoup plus d'importance dans
le piège originel. Elles permettaient d'éclairer la cavité à une époque où l'entrée n'était qu'une mince fente. La même tendance est perceptible dans les
ascidies
juvéniles des plantes de Cephalotus modernes. |
Les crêtes externes garnies de poils sont des émergences peut-être homologues de la saillie des nervures sur la face abaxiale de nombreuses feuilles.
Le Cephalotus est une plante des marais qui exige un sol humide et même mouillé. Cela n'est pas sans poser quelques problèmes quand on sait que le climat du Sud-Ouest de l'Australie est de type méditerranéen.
Les étés y sont secs et chauds et
il ne pleut guère qu'en hiver.
On sait aussi que la plupart des végétaux locaux même ceux des marais ne croissent que l'hiver. Seuls leurs
rhizomes
(p. ex. Byblis gigantea) ou leurs tubercules
(Droséras du sous-genre Ergaleium) survivent à l'été.
Cephalotus follicularis ne possède pas de tels mécanismes. Il serait bien incapable de survivre dans ces conditions s'il ne croissait justement dans une région limitée au climat moins tranché.
| Les journées nuageuses les averses de crachin et une abondante rosée nocturne n'y sont pas rares l'été. Les marais à Cephalotus ne se dessèchent jamais complètement et
restent même carrément mouillés jusque tard dans la saison sèche. On
ne les trouve guère que dans la partie du Sud-Ouest australien où les précipitations annuelles dépassent les 200 cm. |

|

|
Ces régions ont un soubassement granitique produisant par érosion un substrat sablonneux de
pH
4.5. Celui-ci est lessivé par les flux d'eau souterrains en provenance des collines environnantes et qui affleurent ici. Ils emportent les minéraux et empêchent la dégradation des débris végétaux.
Ainsi naissent les marais tourbeux où croît Cephalotus. |
| La zonation y est la suivante : aux pentes plus sèches couvertes de broussailles basses succède une zone humide caractérisée par de petits arbres des broussailles hautes (1.5-3 m) et déjà beaucoup de roseaux.
Le marais proprement dit est trop
mouillé pour les arbres. On n'y trouve que des buissons hauts d'1m-1m50 et des amas disséminés de joncs et d'herbes. |

|

|
C'est parmi ces derniers qu'on trouve communément Cephalotus et aussi dans l'enchevêtrement de racines au pied des buissons ligneux (Beaufortia sparsa
Leptospermum firmum). La zone arborée hantée par les serpents venimeux recèle aussi des Cephalotus. On peut parfois les trouver paradoxalement sur des pentes tourbeuses ou sur les bords surélevés
éventuellement purement sablonneux des marais. |
Ils y croissent en plein soleil associés à des plantes photophiles par exemple Drosera pulchella D. hamiltonii.
Cela n'est possible que là où un flux d'eau affleure. Bien que le sable
y semble sec il suffit d'y enfoncer le doigt pour s'assurer qu'il est mouillé. Dans le marais-même Cephalotus se voit rarement côte à côte avec Drosera hamiltonii qui préfère les endroits ouverts et
Utricularia hookeri plus près de l'eau.
ADAMS R.F. (1978a) Plant parenthood and the single cell. Horticulture 56 n° 10:16-22
(via CPN 7 (197!) n°4:103)
- - (1978b) Cephalotus foIlicularis: the Australian Pitcher Plant.
Am. Hortic. 57 n° 3:4-5
ADAMS R.M. & KOENIGSBERG S.S. & LANGHANS R.W. (1979) In vitro propagation of Cephalotus follicularis. HortScience 14 n° 4:512-513.
ADAMS R.M. & SMITH G.W. (1977) An S.E.M. survey of the five carnivorous pitcher
plant genera. Amer. J. Bot. 64:265-272.
ANDRÉ Ed. (1880) Cephalotus follicularis. L'Illustration horticoie: 117-118 + pl. CCCXCI.
ARBER Agnes (1941) On the morphology of the pitcher leaves in Heliamphora Sarracenia Cephalotus and Nepenthes. Annals of Botany new series 5:563-578.
ARMITAGE David (1984) A beginner's diary of carnivorous plant growing. Carnivorous plant Society Journal (G.B.) Vol. 8-spring:5.
AUBRY René (1985) Cephalotus follicularis. Dionée n°5:11-15.
BAFFRAY M. & BRICE F. & DANTON Ph. & TOURNIER J.-P. (1989) Nature et Culture des Plantes Carnivores. Edisud Aix-en-Provence.
BAILLON H. (1872) Histoire des Plantes III:337 sq. 430 figs. 378-381. Paris Hachette.
- - (1876) Dictionnaire de botanique :700.
BATE-SMITH E.C. (1962) The phenolic constituents of plants and their taxonomic significance. J.
inn. Soc. (Bot.) London 58:95-173.
BELLAIR G. & SAINT-LEGER L. (1900) CéphaIotées. dans: Les Plantes de serre :491-492. (+ rééd. 1939) Librairie des Sciences et des Arts Paris.
BENTHAM (1864) Flora Australiensis II :448 sq. London Reeve
BENTHAM & HOOKER ( ? ) Genera... :655 pl. I
BERGER A. (1930) Crassulaceae. dans: A. ENGLER (éd.) Die Grossen Pflanzenfamilien 18a:352sq.
BOUCHÉ A. (1884) Insectenfressende Pflanzen pl. I
BOWER F.O. (1889) On Dr. MACFARLANE's observations on pitchered insectivorous plants. Annals of
otany Vol. IV n°XIII (Nov. 1889) : 165-168.
BOWMAN Aylene (1985) dans: Plant of the month. Austral. Carniv. Pl. Soc. Bull. Vol. 4 n°1:9-10.
BREHM (1894-96) Merveilles de la Nature: Monde des Plantes I:737.
BROWN Robert (1814a) dans: FLINDERS A voyage to terra australis... in the years 1801-03 (Botanical appendix) I:76 + pl.4; II:601 + pl.4.
BROWN Robert (1814b) General Remarks on the botany of Terra Australis:68 + pl.4.
- - (1832) On the structure and affinities of Cephalotus follicularis. Edinburgh Philosoph. Magaz. :314.
- - (1835) dans: Martius Conspectus n° 178. Réédition de General Remarks on the Botany of Terra Australis: Miscellaneous Botanical Works I (1866):76-78; II (1867):535 sq. ; Atlas II (1868) pl.4. London Hardwicke
CARLQUIST Sherwin (1981) Wood Anatomy of Cephalotaceae. IAWA Bull. I n°4:175-178.
CARR S.G.M. & CARR D.J. (1976) La contribution de la France à la découverte de l'Australie et de sa flore. Endeavour Vol. 35 n°124:21-26.
CHING-I Peng & GOLDBLATT B.A. (1983) Confirmation of the chromosome number in Cephalotaceae and Roridulaceae. Ann. Missouri Bot. Gard. Vol. 70:197-198.
CLEMESHA Steve (1972) dans: News & Views. CPN Vol. 1:54
- - (1973) dans: News & Views. CPN Vol. 2:57.
COTTER Phil (1988) Flowering Cephalotus. New Zealand Carniv. Pl. Soc. J. March 88:9
DAKIN W.J. (1919) The West Australian pitcher plant Cephalotus follicularis and its physiology. J. & Proc. of the Roy. Soc. of W. Australia Vol. 4:37-53.
DE BUHR Larry (1976) Field notes on Cephalotus follicularis in W. Australia. CPN Vol. 5 n° 1:8-9.
DE CANDOLLE A.P. (1825) Prodromus systematis naturalis regni vegetabilis (Dicot.) II:59. Paris TreutteI & Wurst.
DICKSON Alexander (1878) On the structure of the pitcher of Cephalotus follicularis. J. Bot. 2e série VII:1sq + pl.
- - (1881a) On the morphology of the pitcher of Cephalotus follicularis. J. Bot. n.s. X: 129-135.
- - (1881b) [résumé article précédent] Gardeners Chron. VoI. I:373.
- - (1883) [réédition de 1881a] Transact. & Proc. Bot. Soc. Edinb. : 172-181 + pl.V
DIELS L. (1906) Die Pflanzenwelt West Australiens. dans: ENGLER-DRUDE Veget. der Erde VII :357.
- - (1930) Cephalotus follicularis. dans: A. ENGLER Die Grosse Pflanzenfamilien XVIII:71-74.
D.J.R. (1939) Vleeschetende- Planten. dans: Floralia: 173-179.
DUCHARTRE P. (1877) Elêments de Botanique (3e éd.):408-409. Paris Baillière et fils.
DUJARDIN P. (1897) Cephalotus follicularis. Rev. Hortic. Belge XXIII:233-234.
DWYER T. Patrick (1983) Seed structure of carnivorous plants>. CPN Vol. 12 n°1:11.
EICHLER A.W. (1878) Bluetendiagramme construirt und erlautert. II:436. Leipzig Engelmann.
- - (1881) Ueber die Schlauchblaetter von Cephalotus follicularis. Jahrb. des Kgl. Berl. Rot. Gart. I:193-197.
EMBERGER L. (1960) Les végétaux vasculaires. dans: CHADEFAUD M. & EMRERGER L. Traité de Botanique systêmatique II. Paris Masson
ENDLICHER Stéph. (1836) Genera plantarim secundum ordines naturales disposita II:812. Wien Fr. Beck
ENGLER (1919) Saxifragaceae. dans: A. ENGLER (éd.) Das Pflanzenreich IV:117.
ERlCKSON Rika (1968) Plants of Prey. Nedlands W. Australia.
GlLBURT W.H. (1881) Notes on the histology of pitcher plants. Quekett Microsc. J. Vol. 6:151-164
GOEBEL K. (1898-1901) Organographie der Pflanzen (1e éd.):531-532. Jena Froecher
- - (1904) PflanzenphysioIogische Schilderungen II:110-115; 170-173 + pl. III. Marborg Elwertsch
- - (1905) [éd. angl. ] II:337 339.
GREEN Fred (1987) Cephalotus follicularis propagation by leaf cuttings. Carniv. Pl. Soc. (G.B.) Newsletter 1987 n°2:4.
GRIEVE R.J. (1961 ) The genus Cephalotus. Austral. Pl. Vol. I (September):26-28.
HADDON H. (1916) Cephalotus follicularis. The Gardeners' Chronicie 1 April 1916:182.
HAMILTON A.G. (1904) Notes on the W. Australian pitcher plant (Cepalotus follicularis). Proc. Linn. Soc. New S. Wales Vol. 29:36-53.
HANLON Andrew (1983) Growing Cephalotus in Australia. CPN Vol. 12 n°4:95.
HARGRAVE A.L. (1966) My pitcher plant. Austral. Pl. Vol. 4 n°12:37.
HENNERN Holger (1987) Unusual Cephalotus follicularis leaves. Carniv. Pl. Soc. (G.B.) J. Vol. II (spring):4-5.
HOLDCROFT Tony (1989) Cephalotus. Austral. Carniv. Pl. Soc. Bull. Vol. 8 n°4:63-65.
HOOKER W. (1831) Cephalotus follicularis. Curtis's Bot. Mag. LVIII :3118-3119.
HOWELL Fred (1985) dans: Plant of the month. Austr. Carniv. Pl. Soc. Bull. Vol. 4 n°1:7.
IRVING Brian (1988) Control of fungal attack in Cephalotus. New Zeal. Carniv. Pl. Soc. J. December '88.
JAY M. & LEBRETON P. (1972) Recherches chimiotaxinomiques sur les plantes vasculaires: XXVI. Les Flavonoïdes des
Sarracéniacées Népenthacées Droséracées et Céphalotacées. Naturaliste Canadien 99:607-613.
JOHNSON M.A.T. (1979) Chromosome numbers in Akania and Cephalotus. Kew Bull. 34 n°1:37-38.
JUNIPER B.E. & ROBINS R.J. & JOEL D.M. (1989) The Carnivorous Plants. London Academic Press
JUSSIEU (1818) ?
KEIGHERY G.J. (1979) Chromosome counts in Cephalotus. Plant Systematics and Evolution Vol. 133 n°1:103-104.
KERNER Anton von Marilaun (1887) Pflanzenleben I:22 ou 3e éd. (1913) I:315. Adolf Hauser Leipzig & Wien
KERNER & OLIVER (1894) Nat. Hist. of Plants I:131. London Blakkie & Son
KNIGHT A.E. & STEP E. (1905) Living Plants pl.5.
KONDO Katsuhiko (1972) dans: News & Views. CPN Vol. 1:40sq.
KRONFELD (1899) Bilder Atlas zur Pflanzengeographie:170 Leipzig-Berlin
KUSAKABE Isamu (1987) dans: News & Views. CPN Vol. 16 n°4:96.
KUTT Dave (1975) dans: News & Views. CPN Vol. 4:55.
- - (1976) Cultivating Cephalotus follicularis. CPN Vol. 5: 10.
LABILLARDIERE (1800) Novae Hollandiae Plantarum Specimen II:6-7 + pl. 145.
LAMARCK (1823) Encyclopédie méthodique de Botanique t. II Illustr. VI pl .960.
LAWSON TAIT (1878) Transact. Phil. Soc. Birmingham I n°3:125.
LECOUFLE Marcel (1989) Comment choisir et cultiver vos plantes carnivores. Bordas Paris
LEMAIRE Ch. (1847) Cephalotus follicularis. FIore des Serres série 1 n°III pl.290.
LE MAOUT & DECAISNE (1866) Traité général de Botanique:266-267. Paris
LINDLEY John (1835) Key... n°5:45.
- - (1836) Nat. Syst. (2e éd.) :14 London Longmann
- - ( ? ) Veg. Kingdom:428.
LLOYD Francis E. (1932) The carnivorous plants -a review with contribution. Trans. Roy. Soc. Canada III:1-67.
- - (1942) The carnivorous plants. Waltham Mass.
LOWRIE Allen (1978) Cephalotus hunting in the deep South West of Australia. CPN Vol. 7:119sq.
LULLFITZ F. (1960) The W. Australian pitcher plant. Austral. Pl. Vol. 4
N°12:34-35.
MACFARLANE J.M. (1889) Observations on Pitchered Insectivorous PIants part I . Annals of Bot. III:265sq.
- - ( 1893) [idem] part II. Annals of Bot. VII:444sq.
- - (1900) Cephalotus. dans: BAILEY L.H. Standard cyclopaedia Hort. :716.
MACFARLANE J.M. (1910-11) Cephalotaceae. dans: A. ENGLER (éd.) Das PfIanzenreich IV.116.
MANN Phil (1978) dans: News & Views. CPN Vol. 7:35.
- - (1985) The Albany Area. CPN Vol. 14:75sq.
MASTERS M.T. (1869) Vegetable Teratology... :314. London Hardwicke
MAURY. Paul (1887) Note sur l'ascidie du Cephalotus follicularis.
ull. Soc. Hot. de Fr. 34:164-168.
MAZRlMAS Joseph A. (1975) Seed. CPN Vol. 4:45.
- - (1976) On growing the Australian Pitcher Plant. CPN Vol. 5:10sq.
MEISSNER C.F. (1831) Plantarum vascularium genera I:105.
- - (1836-43) [idem] II:74. Lipsiae-Heidmann
MOON Pat (1986) Cephalotus follicularis from seed. Carniv. Pl. Soc. (G.B. ) Newsletter 1986 n°2:5.
- - (1987) Cephalotus seed germination. Carniv. Pl. Soc. (G.B.) Journal Vol. 11(Spring):5.
MORENO Daniel (1988) Boutures de Cephalotus. Dionée n°15:15-17.
MORLEY Stephen (1989) Cephalotus propagation. Carniv. Pl. Soc. (G.B.) Journal Vol. 13(Autumn'89):33.
MÜLLER Jan (1981) Fossil pollen records of extant Angiosperms. Bot. Rev. 47: 1-145.
NEES AB ESENBECK (1844) dans: LEHMANN Planta Preissianae... I:278. Hamburg Simptibus Meissneri
NEGER (1913) Riologie der Pflanzen auf experimenteller Grundlage:331. Stuttgart Ferd. Encke
NICHOLLS K.W. & BOHM B.A. & ORNDUFF R. (1985) Flavanoids and affinities of the Cephalotaceae. Biochem. Syst. Ecol. 13 n°3:261-264.
PARKES D.M. (1980) Adaptative mechanisms of surfaces and glands in some carnivorous plants. M. Sc. thesis Monash Univ. Clayton Victoria Australia
PARKES D.M. & HALLAM N.D. (1984) Adaptations for carnivory in the W. Australian pitcher plant Cephalotus follicularis. Austral. J. Bot. 32:595-604.
PERSCH F. (1981) dans: News & Views. CPM Vol. 19:34.
PIERSON Bruce (1985) dans: Plant of the month. Bull. Austral. Carniv. Pl.
Soc. Vol. 4 n°1:8.
PIETROPAOLO J. & P.A. (1986) Carnivorous Plants of the World. Timber Press Portland Oregon U.S.A.
POIRET (1808) dans: LAMARCK Encyclop. Méthod. Bot. VIII:326.
RAVEN P.H. & AXELROD D.I. (1974) Angiosperm biogeography and past continental movements. Ann. Missouri Bot. Gard. 61:539-673.
RIEDL. Robert (1985) Cephalotus follicularis the W. Australian pitcher plant. CPN Vol. 14:62sq.
ROSE Stephen (1973a) dans: News & Views. CPN Vol. 2:16.
- - (1973b) dans: News & Viexs. CPN VoI. 2:33.
- - (1976) dans: News & Views. CPN Vol. 5:21 .
ROTH Ingrid (1949) Zur Entwicklungsgeschichte des Blattes mit besonderer Beruecksichtigung von Stipular- und Ligularbildungen. PIanta (Berl.) 37:299-336.
- - (1952) Beitraege zur Entwicklungsgeschichte der Schildblaetter. Planta (Berl.) 40:350-376.
SCHNELL Donald E. (1980) Cephalotus follicularis. CPN Vol. 9:50sq.
SCHNIZLEIN A. (1851) Iconogr. III pl. 169.
SCH0LL Bill (1978) dans: News & Views. CPN Vol. 7:99.
SCHWEIGER Josef (1910) Vergleichende Untersuchungen ueber Sarracenia und Cephalotus follicularis betreffs ihren etwaigen systematischen Verwandtschaften. Beihefte Bot. Centralbl. Vol. 25:490-539.
SIBLEY T. (1988) Cephalotus from seed. New ZeaI. Carniv. Pl. Soc. Vol. 6 n°4:10-12.
SLACK A. (1979) Carnivorous Plants.
- - (1986) Insect-eating plants and how to grow them. Sherborne Dorset
SONG Leo C. (1978) dans: Questions and Answers. CPN Vol. 7:59.
SWINBOURNE Rob. F.G. (1985) dans: Plant of the month. Bull. Austral. Carniv. Pl. Soc. Vol. 4 n°1:5-7.
TAKHTAJAN A. (1959) Die Evolution der Angiospermen. Jena
TRATTINICK L. (1812) Arch. I pl.96.
- - (1825) Gen. Nov. Pl. pl .9
TROLL Wilhelm (1932a) Morphologie der Schildfoermigen Blaetter. Planta (Berl. ) 17:153-314.
- - (1932b) Ueber Diplophyllie und verwandte Erscheinungen in der Blattbildung. Planta (Berl.) 15:315-406.
- - (1934) Vergleichende Morphologie der hoeheren Pflanzen. I.2.4 1852-1856; 1866-1877.
VAN DEN HEEDE Ad. (1909) Les plantes carnivores. Revue Hortic. Prat. 11:137. Paris
VINES S.H. (1903) Proc. Linn. Soc. London Oct. 1903.
WAGNER Adolf (1911) Die fleischfressenden Pflanzen.
WARBURG O. (1916) Die Pflanzenwelt II:92. Leipzig & Wien Bibl. Inst.
WARMING (1902) Handb. Syst. Bot. Ed. Moebius:341.
WETTSTEIN R. (1935) Handbuch der systematischen Botanik. Leipzig Deutsche
WHITEHEAD Brian (1973a) dans: News & Views. CPN Vol. 2:4.
- - (1973b) Cephalotus from seed. CPN Vol. 2: 10sq.
WILLIS J.H. (1965) Historical notes on the W. Australian pitcher plant. Western Australian Naturalist 10:1-7.
WINTER K. (1936) Cephalotus follicularis. Gartenflora 85:100-101.
WOOL (1879) Lectures on the Vegetable Kingdom:100.
|
Références anonymes ou incomplètes
|
Album Natuur (1864):58 60-62.
Ann. Hortic. III (1847):397.
Austral. PI. Vol. 8 (1975) n°14:172-173.
Floral Mag. VII (1868) pl.372.
Garden II (1872):340
Garden III (1873):307 401.
Garden XVII (1880):405.
Garden LVII (1900):377.
Gartenflora Deutschlands Russlands und der Schweiz XXV (1876):216.
Gartenflora... 85 (1936):101.
Hort. Reg. II (1833):365.
Jahresb. Niedersachs. Vereins 1-2 (1908-1909):9.
Journ. Hort. LI (1874):327.
Journ. Hort. sér. III XII (1886):95.
Journ. Hort. XXXV (1897):260.
Journ. Hort. LXV (1912):147.
Lyon Horticole 34 (1912):23.
Pareys Blumengartnerei (1931):101.
Paxton Mag. of Botany I (1841):59.
Proc. Roy. Hortic. Soc. 21 (1897-1898):CLVIII-CLIX.
Rhein. Jahrb. Gartenk. & Bot. I (1886):186.
Wien. III. Gart. Zeit. VI (1881):29.
POSTSCRIPTUM SUR L'ÉVOLUTION DES ASCIDIES
|
Lors de l'élaboration de la théorie évolutive présentée dans ce fascicule nous nous étions basés sur les données fournies par TROLL. Dans leurs grands traits celles-ci sont encore acceptées à l'heure actuelle
(voir JUNIPER et al. p. 58)
et ce malgré les critiques d'Ingrid ROTH à la fin des années 40. Cependant nous avons dû mentionner que dans le cas de Cephalotus TROLL s'était trompé sur un point:
la non-fusion
des faisceaux ventraux. En relisant le manuscrit de ce fascicule j'ai été frappé par
un fait qui n'avait pas attiré mon attention jusqu'à présent: la vascularisation de
l'opercule
(p. 52) ne se fait pas par les veines ventrales en tant que telles. Celles-ci se dirigent en réalité vers le
péristome
et ne donnent au couvercle qu'une série de branches. Or si l'on considère les deux moitiés de
l'opercule
comme des folioles fusionnées on s'attendrait à ce que chaque lobe reçoive son faisceau vasculaire propre en d'autres termes que les veines ventrales se
dirigent droit dans
l'opercule ce qui n'est pas le cas. Si l'on aplatit
l'ascidie
reconstituant ainsi les positions relatives des nervures avant la peltation on voit que les lobes de
l'opercule
ne sont que des appendices secondaires. Les faisceaux ventraux de
l'ascidie
retrouvent leur rôle de nervures marginales (suivant le bord de la feuille). On retrouve aussi pour cette dernière une forme
lancéolée
comparable à celle des feuilles d'hiver. Toutefois cela ne nous autorise pas à considérer ces dernières
comme représentant la forme archaïque des
ascidies.
Comme nous l'avons vu la disposition des faisceaux vasculaires
indique clairement la nature tératologique de ces feuilles.
Malgré la fait que vu la différenciation tardive des structures vasculaires la disposition des nervures n'est pas considéré comme un indice important dans la recherche des homologies entre parties des feuilles normales et celles
d'ascidies
la géométrie des faisceaux de
l'opercule
me semble tout de même exclure le modèle foliolaire et sans doute aussi diplophylle. Dans ce dernier cas les nervures des lobes basaux (réfléchis vers le haut) des feuilles
proviennent également des faisceaux les plus latéro-dorsaux ou des ventraux s'il y a tendance à l'unifacialité.
Reste à expliquer la présence occasionnelle d'un lobule operculaire médian dans certaines feuilles tératologiques. Si vous en observez ou si vous connaissez des parallèles de la vascularisation
non-directe de lobules basaux de feuilles faites m'en part!





